
De Darwin a Hug: La historia del Árbol de la Vida
El árbol de la vida es un concepto usado como referencia a las relaciones históricas de parentesco entre los seres vivos. A esto también se le conoce como filogenia, que en biología evolutiva hace caso a nivel de todas las especies existentes y que han existido en el pasado.
Todo se sostiene en que los seres vivos provenimos de otros seres vivos, hasta llegar a LUCA, sigla en inglés del Último Ancestro Común Universal: el primer ser vivo que dio origen a los demás.
La historia del Árbol de la Vida
Nos remontamos a 1837. Específicamente a una de las libretas de Charles Darwin, donde deja expresado que así creía que se podría simbolizar la emergencia de especies. Ya en su primera edición del Origen de las Especies representa de manera más detallada la influencia de la selección natural sobre un género biológico hipotético.
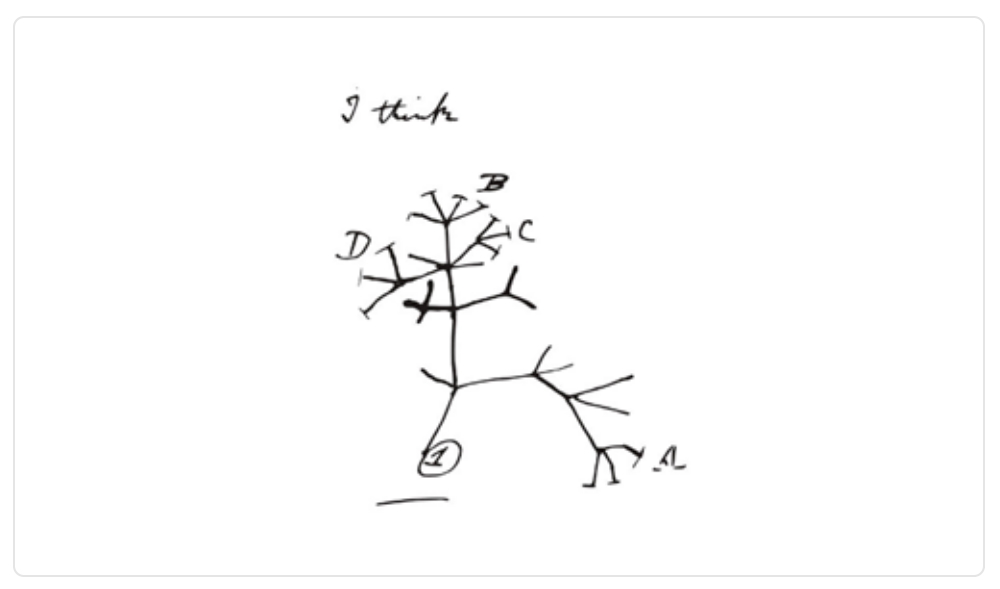
Imagen 1. Dibujo de la representación de la emergencia de especies realizado por Darwin.
Años después, en 1866, Ernest Haeckel propuso árboles de variados grupos de seres vivos. Su árbol de la vida más conocido fue publicado en su libro Morfología General de los Organismos, donde propuso la clasificación de la biota en tres reinos, ampliando la propuesta de Linneo -fundador de la taxonomía- del reino animal y vegetal, agregando los protistas, los cuales son organismos unicelulares y pluricelulares muy básicos.
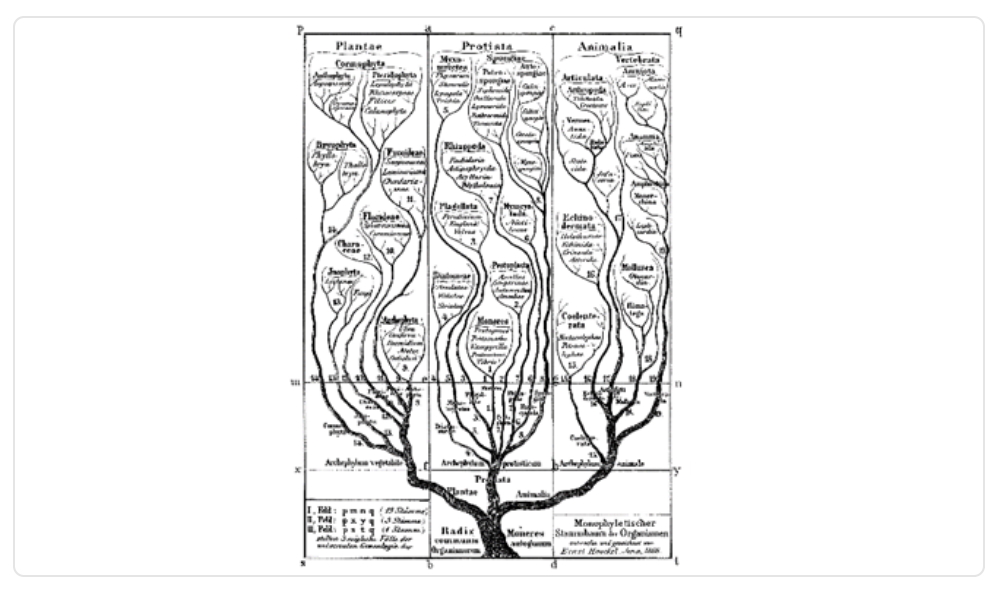
Imagen 2. El primer Árbol de la Vida con tres reinos, propuesto por Haeckel.
Casi un siglo más tarde, en 1956, Herbet Copeland divide el reino protista en monera -exclusivo de bacterias- y protistas -unicelulares y pluricelulares básicos-. Luego, en 1959, Robert Whittaker publica un artículo en donde presenta su nuevo árbol de la vida, el que conservaba los reinos protista, animalia y plantae, pero sumaba a los hongos como un reino aparte basándose en criterios citoquímicos y genéticos por primera vez.
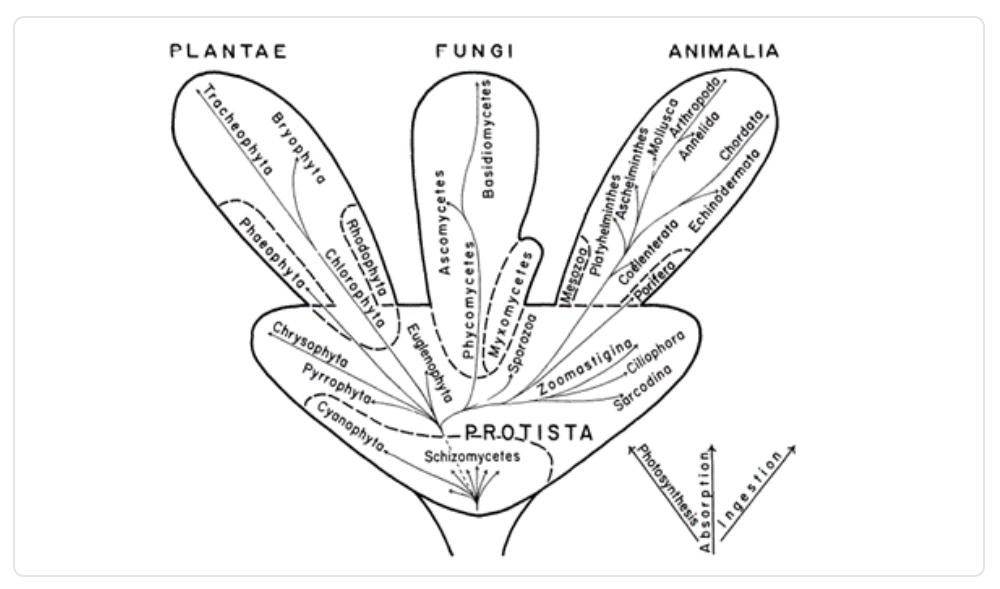
Imagen 3. Clasificación de los seres vivos elaborada por Whittaker.
Diez años después de su primera clasificación, Whittakerdecide hacer una segunda para mostrar a los monera como un reino. Esta versión se queda con un total de cinco reinos: protista, plantae, animalia, monera y hongos. Además, es la que se enseña con más frecuencia en la escuela.
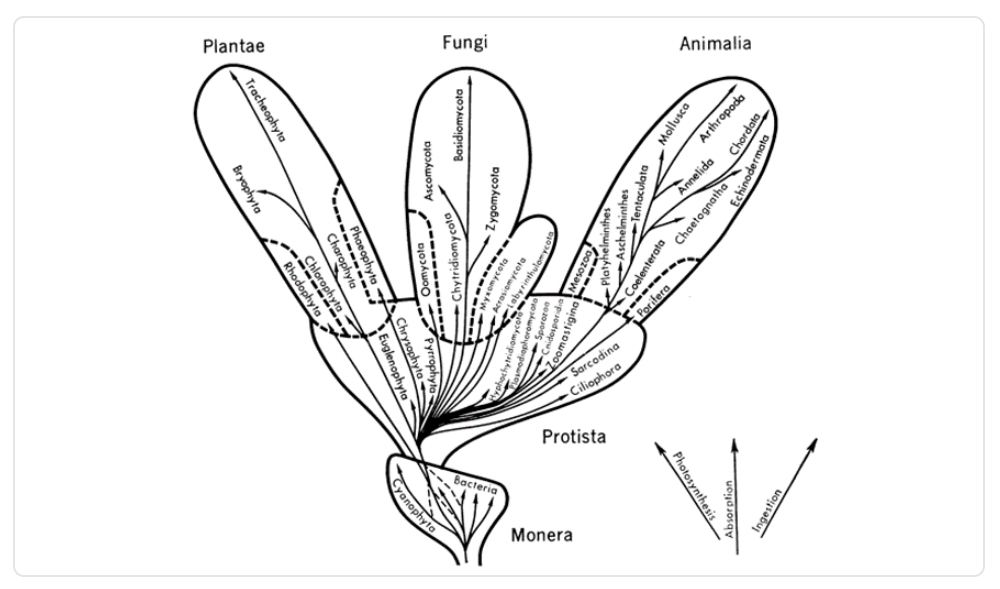
Imagen 4. Propuesta de la segunda clasificación de los reinos y su árbol de la vida, hecha por Whittaker.
La unión hace la fuerza
Con el paso del tiempo, la científica Lynn Margulis, reconocida por su Teoría de la Endosimbiosis Seriada, en 1971 hizo una crítica al árbol de Whittaker y en 1974 propuso su propio árbol con cinco reinos, pero con la categoría adicional de los super reinos: eukaryota y prokariota. Posteriormente, ambos científicos trabajaron en conjunto para mejorar sus propuestas, las cuales fueron presentadas en un artículo en 1978, que no incluía el árbol filogenético.
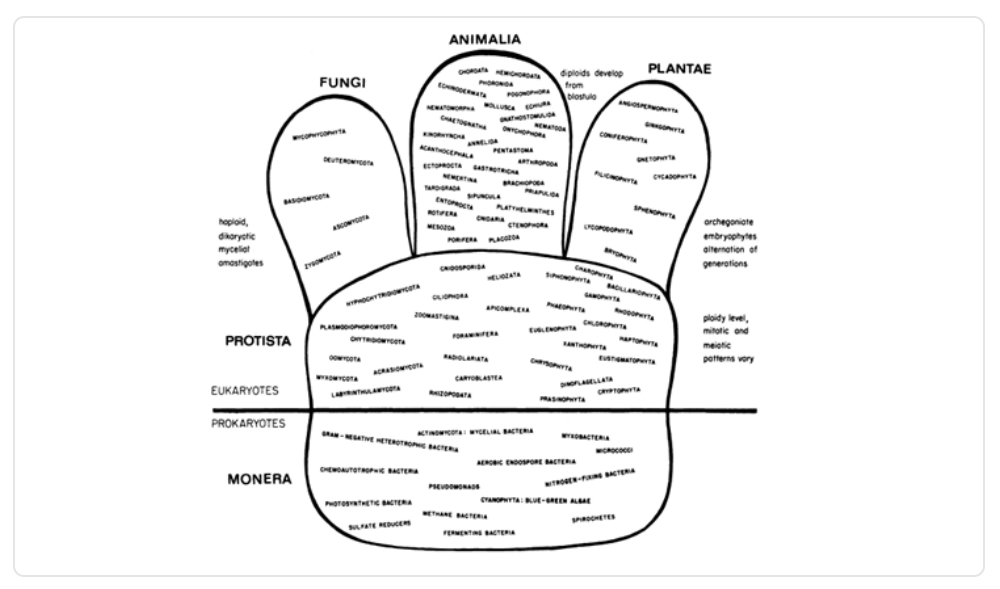
Imagen 5. Filogenia de los cinco reinos en los dos super reinos, propuesta por Margulis.
Siguiendo un trabajo similar, Carl Woese, Otto Kandler y Mark Wheelis realizan en 1990 una nueva propuesta, la cual se basa en clasificar a todos los seres vivos en tres dominios: bacterias, arqueas y eucariotas, todo esto inspirado principalmente en los avances de la genética molecular.
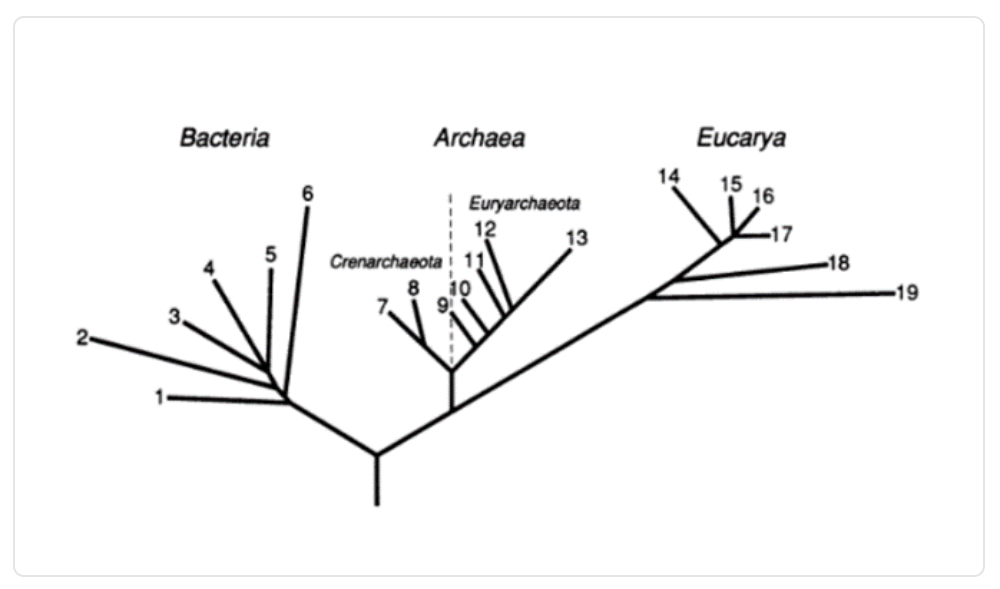
Imagen 6. Representación de los tres dominios realizada por Woese, Kandler y Wheelis.
Es así como vemos que la historia del árbol de la vida ha pasado por diversos criterios y modificaciones. Parte, eso sí, desde un inicio con las características morfológicas, luego los criterios bioquímicos, y posteriormente, los análisis genéticos. Pero ¿por qué sucedieron estos cambios? ¿A qué se debió el avance en esos casos?
¿Por qué quedaron obsoletos los modelos antiguos?
A pesar de que la morfología es uno de los principales criterios de diferenciación y también la base de los primeros árboles de la vida, esta no fue suficiente para separar correctamente a las distintas especies de nuestro planeta.
La razón es simple: si el parentesco se va acercando, observar las discrepancias físicas se hacía mucho más dificultoso. Un claro ejemplo es el caso del jaguar y el guepardo, que se parecen mucho, aunque sean especies diferentes.
Por otro lado, el dimorfismo sexual, basado en dos miembros de una misma especie, pero de distinto sexo, permite examinar diferencias morfológicas notorias, tales como el tamaño, los colores, y formas. Por ende, se hace difícil catalogarlas como miembros de una misma especie si solo nos basamos en sus características morfológicas.
Incluso yendo más lejos, este criterio se vuelve aún más limitado por sí mismo cuando se trata de diferenciar microorganismos -como las bacterias-. Dos individuos se pueden parecer mucho en forma, pero difieren en otras características, catalogándose finalmente como especies distintas.
Por suerte, los principios morfológicos se pueden complementar con características metabólicas para diferenciar distintos tipos de bacterias; en otras palabras, determinar qué utiliza y hace una bacteria para obtener energía. Pero, como sucede con la morfología, si dos bacterias están muy emparentadas, se hace complicado diferenciarlas correctamente.
Otro de los principales problemas que se presentaban en los primeros modelos de árboles de la vida, era que no poseían la capacidad de mostrar la interacción horizontal entre algunos de los grupos filogenéticos que existían.
En aquel entonces, se tenía una concepción vertical de la evolución, es decir, que las nuevas especies aparecían de diferencias entre hijos y padres que con el tiempo terminaban en el surgimiento de nuevas especies.
Aunque cuando nos referimos a microorganismos, esto es diferente. La transferencia de genes y cambios no solo se da de manera vertical, sino que existe un proceso denominado transferencia horizontal de genes. Esta consiste en la capacidad de las bacterias para compartir e intercambiar parte de su genoma entre ellas sin mantener una relación de parentesco.
Nuevo modelo, nuevos fundamentos
Debido a los problemas que presentaban los criterios usados en un inicio, los investigadores y estudiosos de la materia se preguntaron nuevamente: ¿cómo podemos diferenciar a las distintas especies de una manera más precisa y certera?
La respuesta fue encontrada de la mano con el auge del avance científico y tecnológico de la época, donde se planteó la idea de comparar similitudes en el genoma de las especies. En otras palabras, entre más parecido el material genético (ADN) de las especies, más cerca están unas de otras.
Pero esta idea presentaba el problema de que no todos los materiales genéticos son iguales. Para ejemplificarlo, volvemos una vez a las bacterias. Ellas poseen un genoma mucho más pequeño que organismos más complejos, como los humanos.
Así que ¿cómo los comparamos?
A pesar de que todos los materiales genéticos son diferentes, todas las especies provenimos de un único ancestro en común y original, LUCA, lo que nos hace tener algo en común con todos los organismos vivos de nuestro planeta.
También sabemos que hay una estructura que todo ser vivo posee, desde los organismos unicelulares a algunos muy complejos y grandes, como una ballena: los ribosomas.
Entonces, las nuevas relaciones filogenéticas nacen de la comparación genética entre los RNA 16s de todos los seres vivos en nuestro planeta. Este criterio nos otorga una comparación y diferenciación más precisa, además de que nos responde muchas dudas que quedaban con los criterios usados anteriormente.
Por muy morfológica y metabólicamente parecidas que fueran dos especies, si no tenían cierto porcentaje de similitud entre los RNA 16s, se catalogaban como no pertenecientes a la misma especie.
Esta nueva forma de ver las relaciones de parentesco nos abrió los ojos frente a cercanías que antes no habíamos visto, como, por ejemplo, que una orca es más parecida a un delfín que a una ballena por sus genes.
El futuro es hoy
En base a esto, la científica Laura Hug y su equipo propusieron un nuevo árbol de la vida en el 2016. Basado en las similitudes del RNA 16s ribosomal, nos entrega una forma más certera de representar las relaciones de parentesco entre especies, la cual se puede apreciar en la figura a continuación:
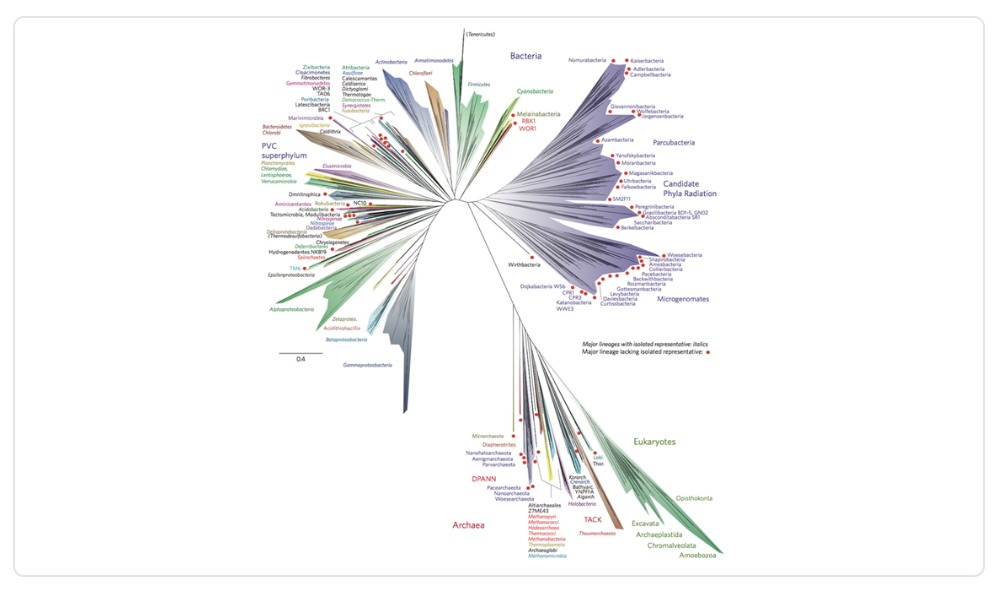
Imagen 7. Nuevo modelo del árbol de la vida(Hug et al., 2016).
Sin embargo, hay que tener en cuenta que estas similitudes o diferencias en el ARN ribosomal 16s no es la vía para clasificar a todas las especies que se van encontrando. Esto es debido a las limitaciones de las técnicas implementadas para la caracterización de las secuencias genéticas, lo que nos genera grandes planteamientos: ¿qué otras cosas nos estamos perdiendo? ¿Qué relaciones no estamos viendo? ¿Cómo será la construcción de los árboles de la vida del futuro?

